Путь воды в растении распадается на три различные по физиологии, строению и протяженности части: по живым клеткам корня; по мертвым элементам ксилемы корня, стебля, черешка и жилок; по живым клеткам листа до испаряющей поверхности.
Большая часть этого пути приходится на долю водопроводящей системы, состоящей из мертвых полых сосудов у покрытосеменных и трахеид у голосеменных растений. У травянистых растений эта часть водного пути достигает десятков сантиметров, а у древесных - многих метров.
Передвижение воды по сосудам ксилемы, длина которых может достигать нескольких десятков сантиметров, происходит довольно легко. Путь через трахеиды более трудный: от одной трахеиды к другой вода проходит через окаймленные поры; очевидно, что движение воды через них испытывает большее сопротивление, чем через сосуды. Измерения показывают, что у лиственных древесных растений проводимость древесины в 3 - 6 раз выше, чем у хвойных. В целом этот путь вода преодолевает куда легче, чем первый и третий через несколько миллиметров или даже долей миллиметров живых клеток - от корневых волосков до сосудов центрального цилиндра и от сосудов, расположенных в жилках листа, до испаряющих клеток мезофилла.
По сосудам и трахеидам вода передвигается, как по полым трубкам, подчиняясь общим гидродинамическим законам, по живым же клеткам корня и листа - осмотическим путем, с помощью разности сосущих сил соседних клеток в правильно возрастающей последовательности. Значительное сопротивление току воды при переходе ее от одной живой клетки к другой делает этот способ совершенно непригодным для передвижения воды на большое расстояние. Поэтому возникновение трахеид у папоротниковидных растений явилось важным этапом в эволюции растительного мира. Еще более совершенной стала водопроводящая система с появлением настоящих сосудов у покрытосеменных растений.
Через растение перекачивается огромное количество воды. С 1 га посевов пшеницы за лето испаряется около 2 тыс. т, клевера - 7,5, капусты - 8 тыс., Т.е. если собрать всю воду, расходуемую 1 га клевера или капусты, то получится водный бассейн площадью 1 га и глубиной 75- 80 см. Еловые молодняки южной тайги за год тратят 4,5 тыс. т воды с 1 га лесной площади, сосновые - 5,0, ольховые - до 11 тыс. т.
Видно, что леса расходуют не меньше, а даже больше, чем некоторые сельскохозяйственные культуры. эти колоссальные расходы восполняются за счет деятельности корневых систем, с достаточной быстротой всасывающих воду из почвы. На этом основана осушающая роль леса в условиях заболачивающихся лесных почв. Поддержанию водного баланса служат и хорошо развитая проводящая воду система, без задержки подающая воду к листьям, а также наличие покровных тканей, защищающих растение от излишней потери воды.
Каковы те силы, которые осуществляют непрерывный ток воды от корней через стебель и листья? У травянистых низкорослых растений механизм перетекания ксилемного сока понять легко. Корневое давление нагнетает воду в сосуды центрального цилиндра корня, а сосущие силы, возникающие в листьях благодаря процессу транспирации, притягивают эту воду. Тем самым создается постоянный ток воды по всему растению.
Процесс подъема воды от корней до листьев носит название восходящего тока, в отличие от нисходящего тока органических веществ от листьев к корням. Корневое давление, создающееся благодаря метаболизму корневых окончаний, получило название нижнего концевого двигателя водного тока. Притягивающие же воду сосущие силы листьев называют верхним концевым двигателем водного тока.
Труднее объяснить непрерывность водного столба у гигантов растительного мира - эвкалиптов, секвой и некоторых других древесных растений, высота которых достигает 140 м. Наши обычные деревья также имеют довольно большие размеры: береза - до 25 м, дуб - 40 м, сосна и ель - до 50 м. К этому следует добавить и значительную протяженность водопроводящей системы корней.
Водный ток испытывает и преодолевает силу земного притяжения, силу тяжести. В силу этого, например, обычные поршневые насосы не могут поднять воду с глубины более 10 м, ибо этот 10-метровый столб воды соответствует давлению в 1 атм. Кроме того, движение воды по ксилеме испытывает довольно значительное сопротивление, особенно у представителей голосеменных древесных растений.
Объяснение того, что водный столб протяженностью многие десятки метров не разрывается, находим в теории сцепления (когезии) и смачивания стенок сосудов и трахеид водой (адгезии). Действительно, между молекулами передвигающейся воды существуют значительные силы сцепления, заставляющие эти молекулы следовать друг за другом. Этому способствует и то, что водопроводящие элементы представляют как бы единое целое с водным потоком, так как стенки их полностью смочены, насыщены водой. В них нет воздуха. В таком состоянии они оказывают минимальное сопротивление движущемуся потоку. Кроме того, само строение сосудов не способствует передвижению пузырьков воздуха из одного сосуда в другой.
Все это весьма сильно отличает условия, создающиеся в дереве, от условий в поршневых насосах. В последних между стенками цилиндра и поршня постоянно появляются пузырьки воздуха, нарушающие целостность водного столба. Происходит обрыв этого столба при подъеме на высоту более 10 м.
Для того чтобы поднять воду на высоту 100 м, необходимо наличие сосущих сил в кроне дерева порядка 30 - 35 атм: на преодоление силы тяжести - 10 атм, сопротивления фильтрации через поперечные стенки сосудов - 20 - 25 атм. В природной обстановке леса такие величины часто регистрируются экспериментально. Поэтому с чисто физической точки зрения представляется возможным объяснить подъем воды на высоту 100 м и более.
В процесс е транспирации в листьях деревьев возникают сосущие силы, достигающие десятков атмосфер. Листья насасывают воду из стебля, вследствие чего в сосудах возникает отрицательное давление разрежение. Такое состояние можно наблюдать при помощи несложных приборов: в теплый летний день при интенсивной потере воды стволы деревьев уменьшаются в диаметре. Другой способ заключается в том, что при срезании интенсивно транспирирующей ветки в подкрашенной воде отмечается мгновенное проникновение краски через поверхность среза благодаря расширению сосудов.
В зависимости от анатомического строения древесины линейная скорость восходящего тока колеблется от 1 - 6 м/ч у хвойных и рассеянно-сосудистых древесных пород до 25 - 60 м/ч у кольцесосудистых. Такая скорость зарегистрирована летом в полдень. Скорость передвижения воды по дереву в течение суток изменяется и в основном соответствует интенсивности транспирации. Существует и светозависимый восходящий водный поток в растениях, тесно не связанный с транспирационной активностью (В.Г. Реуцкий).
В самом дереве быстрее всего вода передвигается в стволе и медленнее - в наиболее молодых ветвях. Среднее положение по этому показателю занимают старые ветви.
Особенности водного тока по стволу дерева:
· С помощью изотопной техники и введения в ствол красок было показано, что у большинства древесных растений водный ток в стволе передвигается по спирали. Это тесно связано с макростроением древесного ствола, что затрудняет ответ на вопрос, какая часть корневой системы питает водой ту или иную сторону кроны дерева.
· Передвижение воды в радиальном направлении осуществляется медленнее и происходит через поры на стенках сосудов и трахеид. Оно имеет значение в поддержании нормальной оводненности живых элементов древесины и коры.
· Неравномерность водного тока. Далеко не вся древесина служит местом проведения воды. У ядровых древесных растений (сосна, дуб) для этого служит только заболонь. При этом более активны в проведении воды последние годичные слои древесины. Это объясняется тем, что только эти слои переходят в древесину однолетних ветвей, тесно связанную с водопроводящей системой листьев. У хвойных, в частности у ели, проводящие пучки хвои сообщаются, по-видимому, с несколькими годичными кольцами. Старые годичные кольца просто не достигают кроны, они выклиниваются по мере увеличения высоты дерева. У ряда древесных пород (акация белая, фисташка, ясень) вода проводится всего лишь 1 - 3 последними годичными слоями заболони. Такая же картина наблюдается и у заболонных, и у спелодревесных пород (осина, береза, липа), но у них число годичных слоев, проводящих воду, несколько больше.
· Можно назвать и такую особенность восходящего тока в стволе дерева, как его изолированность. В общих чертах она присуща и травянистым растениям. Тем не менее, у древесных растений она выражена в большей степени. Проводящая воду древесина ствола отделена от внешней среды не только живыми клетками камбия и флоэмы, но и толстой пробкой или коркой, ограничивающих связь древесины с воздухом атмосферы.
· Такое свойство восходящего тока, как обратuмость, иногда наблюдается в природе у ряда древесных пород, в том числе у некоторых хвойных (ели, пихты), при укоренении нижних ветвей, соприкасающихся с почвой, или верхушек у вываленных деревьев. Ток воды здесь идет в обратном направлении - от морфологически верхнего конца к морфологически нижнему концу.
Восходящий ток обеспечивает все живые клетки растения водой и минеральными элементами. Очень важна роль восходящего тока и в обеспечении живых клеток корней, ствола и ветвей кислородом, ибо проницаемость тканей коры, камбия, древесины для газов весьма низка. Этот растворенный в воде восходящего тока кислород идет на осуществление процесса дыхания живых элементов дерева.
Восходящий ток влияет и на оводненность тканей, в частности древесины ствола. В молодом возрасте до образования ядра наибольшей влажностью обладает древесина хвойных пород. Это различие, очевидно, связано с отсутствием у них по сравнению с лиственными породами либриформа.
Резкие изменения оводненности древесины наблюдаются течение года. Так, у хвойных древесных растений самая низкая влажность отмечается в летние месяцы, а самая высокая - зимой. В поздневесеннее и раннеосеннее время влажность древесины занимает среднее положение. Влажность ядровой древесины практически остается неизменной и самой низкой. У лиственных древесных пород отмечено два периода пониженной влажности - летний и во второй половине зимы и два повышенной - весенний во время сокодвижения и зимний - в первой половине зимы.
Влажность древесины молодых деревьев несколько выше, а амплитуда колебаний ее больше, чем у старых деревьев. Содержание воды в древесине меняется и в течение летних суток: наиболее высокое рано утром, а низкое - в полдень.
Вода, запасенная в середине ствола, способна передвигаться в молодые побеги, что особенно важно при отсутствии доступной для растений воды в почве (засуха, морозы). Например, 100- летняя сосна может переносить засушливый период за счет внутренних запасов воды в стволе в течение целого месяца.
Перетекающая по растению вода отличается от метаболической воды, которая непосредственно используется в различных процессах обмена веществ. Вода постоянно обменивается в клетках растений. С помощью современной техники, в частности изотопной, удалось показать, что самый быстрый обмен внутритканевой воды на внешнюю воду происходит в корнях растений, а самый медленный - в стеблях. Промежуточное положение занимают листья.
Похожая информация.
Вода, поглощенная клетками корня, под влиянием разности водных потенциалов, которые возникают благодаря транспирации, а также силе корневого давления, передвигается до проводящих путей ксилемы. Согласно современным представлениям, вода в корневой системе передвигается не только по живым клеткам. Еще в 1932 г. немецкий физиолог Мюнх развил представление о существовании в корневой системе двух относительно не зависимых друг от друга объемов, по которым передвигается вода, - апопласта и симпласта. Апопласт – это свободное пространство корня, в которое входят межклетные промежутки, оболочки клеток, а также сосуды ксилемы. Симпласт – это совокупность протопластов всех клеток, отграниченных полупроницаемой мембраной. Благодаря многочисленным плазмодесмам, соединяющим между собой протопласт отдельных клеток, симпласт представляет единую систему. Апопласт, по-видимому, не непрерывен, а разделен на два объема. Первая часть апопласта расположена в коре корня до клеток эндодермы, вторая – по другую сторону клеток эндодермы, и включает в себя сосуды ксилемы. Клетки эндодермы благодаря пояскам Каспари представляют как бы барьер для передвижения воды по свободному пространству (межклетникам и клеточным оболочкам). Для того чтоб попасть в сосуды ксилемы, вода должна пройти через полупроницаемую мембрану и главным образом по апопласту и лишь частично по симпласту. Однако в клетках эндодермы передвижение воды идет, по-видимому, по симпласту. Далее вода поступает в сосуды ксилемы. Затем передвижение воды идет по сосудистой системе корня, стебля и листа.
Из сосудов стебля вода движется через черешок или листовое влагалище в лист. В листовой пластинке водопроводящие сосуды расположены в жилках. Жилки, постепенно разветвляясь, становятся более мелкими. Чем гуще сеть жилок, тем меньшее сопротивление встречает вода при передвижении к клеткам мезофилла листа. Иногда мелких ответвлений жилок листа так много, что они подводят воду почти к каждой клетке. Вся вода в клетке находится в равновесном состоянии. Иначе говоря, в смысле насыщенности водой, имеется равновесие между вакуолью, цитоплазмой и клеточной оболочкой, их водные потенциалы равны. Вода передвигается от клетки к клетке благодаря градиенту сосущей силы.
Вся вода в растении представляет единую взаимосвязанную систему. Поскольку между молекулами воды имеются силы сцепления (когезия), вода поднимается на высоту значительно большую 10 м. сила сцепления увеличивается, так как молекулы воды обладают большим сродством друг к другу. Силы сцепления обладают и между водой и стенками сосудов.
Степень натяжения водных нитей в сосудах зависит от соотношения процессов поглощения и испарения воды. Все это позволяет растительному организму поддерживать единую водную систему и не обязательно восполнять каждую каплю испаряемой воды.
В том случае, если в отдельные членики сосудов попадает воздух, они, по-видимому, выключается из общего тока проведения воды. Таков путь передвижения воды по растению (рис. 2).
Рис.2. Путь воды в растении.
Скорость перемещения воды по растению в течение суток изменяется. В дневные часы она на много больше. При этом разные виды растений различаются по скорости передвижения воды. Изменение температуры, введение метаболических ингибиторов не влияют на передвижение воды. Вместе с тем этот процесс, как и следовало ожидать, очень сильно зависит от скорости транспирации и от диаметра водопроводящих сосудов. В более широких сосудах вода встречает меньшее сопротивление. Однако надо учитывать, что в более широкие сосуды могут попасть пузырьки воздуха или произойти какие-либо иные нарушения тока воды.
Филогенетически корень возник позже стебля и листа - в связи с переходом растений к жизни на суше и вероятно, произошёл от корнеподобных подземных веточек. У корня нет ни листьев, ни в определённом порядке расположенных почек. Для него характерен верхушечный рост в длину, боковые разветвления его возникают из внутренних тканей, точка роста покрыта корневым чехликом. Корневая система формируется на протяжении всей жизни растительного организма. Иногда корень может служить местом отложения в запас питательных веществ. В таком случае он видоизменяется.
Виды корней
Главный корень образуется из зародышевого корешка при прорастании семени. От него отходят боковые корни.
Придаточные корни развиваются на стеблях и листьях.
Боковые корни представляют собой ответвления любых корней.
Каждый корень (главный, боковые, придаточные) обладает способностью к ветвлению, что значительно увеличивает поверхность корневой системы, а это способствует лучшему укреплению растения в почве и улучшению его питания.
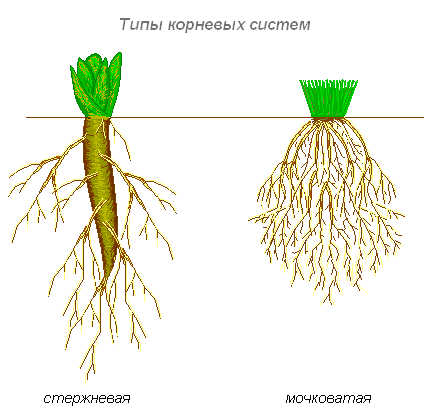
Типы корневых систем
Различают два основных типа корневых систем: стержневая, имеющая хорошо развитый главный корень, и мочковатая. Мочковатая корневая система состоит из большого числа придаточных корней, одинаковых по величине. Вся масса корней состоит из боковых или придаточных корешков и имеет вид мочки.
Сильно разветвлённая корневая система образует огромную поглощающую поверхность. Например,
- общая длина корней озимой ржи достигает 600 км;
- длина корневых волосков — 10 000 км;
- общая поверхность корней — 200 м 2 .
Это во много раз превышает площадь надземной массы.
Если у растения хорошо выражен главный корень и развиваются придаточные корни, то формируется корневая система смешанного типа (капуста, помидор).
Внешнее строение корня. Внутреннее строение корня
Зоны корня

Корневой чехлик
Корень растёт в длину своей верхушкой, где находятся молодые клетки образовательной ткани. Растущая часть покрыта корневым чехликом, защищающим кончик корня от повреждений, и облегчает продвижение корня в почве во время роста. Последняя функция осуществляется благодаря свойству внешних стенок корневого чехлика покрываться слизью, что уменьшает трение между корнем и частичками почвы. Могут даже раздвигать частички почвы. Клетки корневого чехлика живые, часто содержат зёрна крахмала. Клетки чехлика постоянно обновляются за счёт деления. Участвует в положительных геотропических реакциях (направление роста корня к центру Земли).

Клетки зоны деления активно делятся, протяженность этой зоны у разных видов и у разных корней одного и того же растения неодинакова.
За зоной деления расположена зона растяжения (зона роста). Протяжённость этой зоны не превышает нескольких миллиметров.
По мере завершения линейного роста наступает третий этап формирования корня — его дифференциация, образуется зона дифференциации и специализации клеток (или зона корневых волосков и всасывания). В этой зоне уже различают наружный слой эпиблемы (ризодермы) с корневыми волосками, слой первичной коры и центральный цилиндр.
Строение корневого волоска
Корневые волоски — это сильно удлинённые выросты наружных клеток, покрывающих корень. Количество корневых волосков очень велико (на 1 мм 2 от 200 до 300 волосков). Их длина достигает 10 мм. Формируются волоски очень быстро (у молодых сеянцев яблони за 30-40 часов). Корневые волоски недолговечны. Они отмирают через 10-20 дней, а на молодой части корня отрастают новые. Это обеспечивает освоение корнем новых почвенных горизонтов. Корень непрерывно растёт, образуя всё новые и новые участки корневых волосков. Волоски могут не только поглощать готовые растворы веществ, но и способствовать растворению некоторых веществ почвы, а затем всасывать их. Участок корня, где корневые волоски отмерли, некоторое время способен всасывать воду, но затем покрывается пробкой и теряет эту способность.

Оболочка волоска очень тонкая, что облегчает поглощение питательных веществ. Почти всю клетку волоска занимает вакуоль, окружённая тонким слоем цитоплазмы. Ядро находится в верхней части клетки. Вокруг клетки образуется слизистый чехол, который содействует склеиванию корневых волосков с частицами почвы, что улучшает их контакт и повышает гидрофильность системы. Поглощению способствует выделение корневыми волосками кислот (угольной, яблочной, лимонной), которые растворяют минеральные соли.
Корневые волоски играют и механическую роль — они служат опорой верхушке корня, которая проходит между частичками почвы.
Под микроскопом на поперечном срезе корня в зоне всасывания видно его строение на клеточном и тканевом уровнях. На поверхности корня — ризодерма, под ней — кора. Наружный слой коры — экзодерма, вовнутрь от неё — основная паренхима. Её тонкостенные живые клетки выполняют запасающую функцию, проводят растворы питательных веществ в радиальном направлении — от всасывающей ткани к сосудам древесины. В них же происходит синтез ряда жизненно важных для растения органических веществ. Внутренний слой коры — эндодерма. Растворы питательных веществ, поступающие из коры в центральный цилиндр через клетки эндодермы, проходят только через протопласт клеток.

Кора окружает центральный цилиндр корня. Она граничит со слоем клеток, долго сохраняющих способность к делению. Это перицикл. Клетки перицикла дают начало боковым корням, придаточным почкам и вторичным образовательным тканям. Вовнутрь от перицикла, в центре корня, находятся проводящие ткани: луб и древесина. Вместе они образуют радиальный проводящий пучок.
Проводящая система корня проводит воду и минеральные вещества из корня в стебель (восходящий ток) и органические вещества из стебля в корень (нисходящий ток). Состоит она из сосудисто-волокнистых пучков. Основными слагаемыми частями пучка являются участки флоэмы (по ним вещества передвигаются к корню) и ксилемы (по которым вещества передвигаются от корня). Основные проводящие элементы флоэмы — ситовидные трубки, ксилемы — трахеи (сосуды) и трахеиды.
Процессы жизнедеятельности корня
Транспорт воды в корне
Всасывание воды корневыми волосками из почвенного питательного раствора и проведение её в радиальном направлении по клеткам первичной коры через пропускные клетки в эндодерме к ксилеме радиального проводящего пучка. Интенсивность поглощения воды корневыми волосками называется сосущей силой (S), она равна разнице между осмотическим (P) и тургорным (T) давлением: S=P-T.
Когда осмотическое давление равно тургорному (P=T), то S=0, вода перестаёт поступать в клетку корневого волоска. Если концентрация веществ почвенного питательного раствора будет выше, чем внутри клетки, то вода будет выходить из клеток и наступит плазмолиз — растения завянут. Такое явление наблюдается в условиях сухости почвы, а также при неумеренном внесении минеральных удобрений. Внутри клеток корня сосущая сила корня возрастает от ризодермы по направлению к центральному цилиндру, поэтому вода движется по градиенту концентрации (т. е. из места с большей её концентрацией в место с меньшей концентрацией) и создаёт корневое давление, которое поднимает столбик воды по сосудам ксилемы, образуя восходящий ток. Это можно обнаружить на весенних безлистных стволах, когда собирают «сок», или на срезанных пнях. Истекание воды из древесины, свежих пней, листьев, называется «плачем» растений. Когда распускаются листья, то они тоже создают сосущую силу и притягивают воду к себе — образуется непрерывный столбик воды в каждом сосуде — капиллярное натяжение. Корневое давление является нижним двигателем водного тока, а сосущая сила листьев — верхним. Подтвердить это можно с помощью несложных опытов.
Всасывание воды корнями
Цель: выяснить основную функцию корня.
Что делаем: растение, выращенное на влажных опилках, отряхнём его корневую систему и опустим в стакан с водой его корни. Поверх воды для защиты её от испарения нальём тонкий слой растительного масла и отметим уровень.

Что наблюдаем: через день-два вода в ёмкости опустилась ниже отметки.

Результат: следовательно, корни всосали воду и подали её наверх к листьям.
Можно ещё проделать один опыт, доказывающий всасывание питательных веществ корнем.
Что делаем: срежем у растения стебель оставив пенёк высотой 2-3 см. На пенёк наденем резиновую трубку длиной 3 см, а на верхний конец наденем изогнутую стеклянную трубку высотой 20-25 см.
Что наблюдаем: вода в стеклянной трубке поднимается, и вытекает наружу.
Результат: это доказывает, что воду из почвы корень всасывает в стебель.

А влияет ли температура воды на интенсивность всасывания корнем воды?
Цель: выяснить, как температура влияет на работу корня.
Что делаем: один стакан должен быть с тёплой водой (+17-18ºС), а другой с холодной (+1-2ºС).
Что наблюдаем: в первом случае вода выделяется обильно, во втором — мало, или совсем приостанавливается.
Результат: это является доказательством того, что температура сильно влияет на работу корня.

Тёплая вода активно поглощается корнями. Корневое давление повышается.

Холодная вода плохо поглощается корнями. В этом случае корневое давление падает.
Минеральное питание
Физиологическая роль минеральных веществ очень велика. Они являются основой для синтеза органических соединений, а также факторами, которые изменяют физическое состояние коллоидов, т.е. непосредственно влияют на обмен веществ и строение протопласта; выполняют функцию катализаторов биохимических реакций; воздействуют на тургор клетки и проницаемость протоплазмы; являются центрами электрических и радиоактивных явлений в растительных организмах.
Установлено, что нормальное развитие растений возможно только при наличии в питательном растворе трёх неметаллов — азота, фосфора и серы и — и четырёх металлов — калия, магния, кальция и железа. Каждый из этих элементов имеет индивидуальное значение и не может быть заменён другим. Это макроэлементы, их концентрация в растении составляет 10 -2 –10%. Для нормального развития растений нужны микроэлементы, концентрация которых в клетке составляет 10 -5 –10 -3 %. Это бор, кобальт, медь, цинк, марганец, молибден др. Все эти элементы есть в почве, но иногда в недостаточном количестве. Поэтому в почву вносят минеральные и органические удобрения.
Растение нормально растёт и развивается в том случае, если в окружающей корни среде будут содержаться все необходимые питательные вещества. Такой средой для большинства растений является почва.
Дыхание корней
Для нормального роста и развития растения необходимо чтобы к корню поступал свежий воздух. Проверим, так ли это?
Цель: нужен ли воздух корню?
Что делаем: возьмём два одинаковых сосуда с водой. В каждый сосуд поместим развивающие проростки. Воду в одном из сосудов каждый день насыщаем воздухом с помощью пульверизатора. На поверхность воды во втором сосуде нальём тонкий слой растительного масла, так как оно задерживает поступление воздуха в воду.

Что наблюдаем: через некоторое время растение во втором сосуде перестанет расти, зачахнет, и в конце концов погибнет.

Результат: гибель растения наступает из-за недостатка воздуха, необходимого для дыхания корня.
Видоизменения корней
У некоторых растений в корнях откладываются запасные питательные вещества. В них накапливаются углеводы, минеральные соли, витамины и другие вещества. Такие корни сильно разрастаются в толщину и приобретают необычный внешний вид. В формировании корнеплодов участвуют и корень, и стебель.
Корнеплоды
Если запасные вещества накапливаются в главном корне и в основании стебля главного побега, образуются корнеплоды (морковь). Растения, образующие корнеплоды, в основном двулетники. В первый год жизни они не цветут и накапливают в корнеплодах много питательных веществ. На второй — они быстро зацветают, используя накопленные питательные вещества и образуют плоды и семена.
Корневые клубни

У георгина запасные вещества накапливаются в придаточных корнях, образуя корневые клубни.
Бактериальные клубеньки
Своеобразно изменены боковые корни у клевера, люпина, люцерны. В молодых боковых корешках поселяются бактерии, что способствует усвоению газообразного азота почвенного воздуха. Такие корни приобретают вид клубеньков. Благодаря этим бактериям эти растения способны жить на бедных азотом почвах и делать их более плодородными.
Ходульные

У пандуса, произрастающего в приливно-отливной зоне, развиваются ходульные корни. Они высоко над водой удерживают на зыбком илистом грунте крупные облиственные побеги.
Воздушные

У тропических растений, живущих на ветвях деревьев, развиваются воздушные корни. Они часто встречаются у орхидей, бромелиевых, у некоторых папоротников. Воздушные корни свободно висят в воздухе, не достигая земли и поглощая попадающую на них влагу от дождя или росы.
Втягивающие

У луковичных и клубнелуковичных растений, например у крокусов, среди многочисленных нитевидных корней имеется несколько более толстых, так называемых втягивающих, корней. Сокращаясь, такие корни втягивают клубнелуковицу глубже в почву.
Столбовидные

У фикуса развиваются столбовидные надземные корни, или корни-подпорки.
Почва как среда обитания корней
Почва для растений является средой, из которой оно получает воду и элементы питания. Количество минеральных веществ в почве зависит от специфических особенностей материнской горной породы, деятельности организмов, от жизнедеятельности самих растений, от типа почвы.
Почвенные частицы конкурируют с корнями за влагу, удерживая её своей поверхностью. Это так называемая связанная вода, которая подразделяется на гигроскопическую и плёночную. Удерживается она силами молекулярного притяжения. Доступная растению влага представлена капиллярной водой, которая сосредоточена в мелких порах почвы.
Между влагой и воздушной фазой почвы складываются антагонистические отношения. Чем больше в почве крупных пор, тем лучше газовый режим этих почв, тем меньше влаги удерживает почва. Наиболее благоприятный водно-воздушный режим поддерживается в структурных почвах, где вода и воздух находятся одновременно и не мешают друг другу — вода заполняет капилляры внутри структурных агрегатов, а воздух — крупные поры между ними.
Характер взаимодействия растения и почвы в значительной степени связан с поглотительной способностью почвы — способностью удерживать или связывать химические соединения.
Микрофлора почвы разлагает органические вещества до более простых соединений, участвует в формировании структуры почвы. Характер этих процессов зависит от типа почвы, химического состава растительных остатков, физиологических свойств микроорганизмов и других факторов. В формировании структуры почвы принимают участие почвенные животные: кольчатые черви, личинки насекомых и др.
В результате совокупности биологических и химических процессов в почве образуется сложный комплекс органических веществ, который объединяют термином «гумус».
Метод водных культур
В каких солях нуждается растение, и какое влияние оказывают они на рост и развитие его, было установлено на опыте с водными культурами. Метод водных культур — это выращивание растений не в почве, а в водном растворе минеральных солей. В зависимости от поставленной цели в опыте можно исключить отдельную соль из раствора, уменьшить или увеличить ее содержание. Было выяснено, что удобрения, содержащие азот, способствуют росту растений, содержащие фосфор — скорейшему созреванию плодов, а содержащие калий — быстрейшему оттоку органических веществ от листьев к корням. В связи с этим содержащие азот удобрения рекомендуется вносить перед посевом или в первой половине лета, содержащие фосфор и калий — во второй половине лета.
С помощью метода водных культур удалось установить не только потребность растения в макроэлементах, но и выяснить роль различных микроэлементов.
В настоящее время известны случаи, когда выращивают растения методами гидропоники и аэропоники.
Гидропоника — выращивание растений в сосудах, заполненных гравием. Питательный раствор, содержащий необходимые элементы, подаётся в сосуды снизу.
Аэропоника — это воздушная культура растений. При этом способе корневая система находится в воздухе и автоматически (несколько раз в течение часа) опрыскивается слабым раствором питательных солей.
Вода, поглощенная клетками корня, под влиянием разности водных потенциалов, которые возникают благодаря транспирации, а также силе корневого давления, передвигается до проводящих элементов ксилемы. Согласно современным представлениям, вода в корневой системе передвигается не только по живым клеткам. В корневой системе существуют два относительно независимых друг от друга объема, по которым передвигается вода,- апопласт и симпласта. Апопласт - это свободное пространство корня, в которое входят межклетные промежутки, оболочки клеток, а также сосуды ксилемы. Симпласт - это совокупность протопластов всех клеток, отграниченных полупроницаемой мембраной. Благодаря многочисленным плазмодесмам, соединяющим между собой протопласт отдельных клеток, симпласт представляет единую систему. Апопласт, по-видимому, не непрерывен, а разделен на два объема. Первая часть апопласта расположена в коре корня до клеток эндодермы, вторая - по другую сторону клеток эндодермы, и включает в себя сосуды ксилемы. Клетки эндодермы благодаря пояскам Каспари представляют как бы барьер для передвижения воды по свободному пространству (межклетникам и клеточным оболочкам). Для того чтобы попасть в сосуды ксилемы, вода должна пройти через полупроницаемую мембрану и протоплазму клеток эндодермы. Передвижение воды по коре корпя идет главным образом по апопласту и лишь частично по симпласту. Однако в клетках эндодермы передвижение воды идет, по-видимому, по симпласту. Далее вода поступает в сосуды ксилемы. Это, очевидно, вызвано противоположными изменениями в процессах обмена с разных сторон клеток. Согласно другой, это следствие секреции солей в сосуды ксилемы, в результате чего там создается повышенное осмотическое давление. Дальнейшее передвижение воды идет по сосудистой системе корня, стебля и листа. Проводящие элементы ксилемы состоят из сосудов (трахей) и трахеид.
Опыты с кольцеванием показали, что восходящий ток воды по растению движется в основном по ксилеме. В сосудах ксилемы вода встречает незначительное сопротивление, что, естественно, облегчает передвижение воды на большие расстояния. Правда, в настоящее время признается, что некоторое количество воды передвигается и вне сосудистой системы. Однако по сравнению с ксилемой сопротивление движению воды других тканей значительно больше (не менее чем на три порядка). Это приводит к тому, что вне ксилемы движется всего от 1 до 10% общего потока воды.
Из сосудов стебля вода попадает в сосуды листа. Вода движется из стебля через черешок или листовое влагалище в лист. В листовой пластинке водопроводящие сосуды расположены в жилках. Жилки, постепенно разветвляясь, становятся все более мелкими. Чем гуще сеть жилок, тем меньшее сопротивление встречает вода при передвижении к клеткам мезофилла листа. Именно поэтому густота жилкования листа считается одним из важнейших признаков ксероморфной структуры - отличительной чертой растений, устойчивых к засухе.
Иногда мелких ответвлений жилок листа так много, что они подводят воду почти к каждой клетке. Вся вода в клетке находится в равновесном состоянии. Иначе говоря, в смысле насыщенности водой, имеется равновесие между вакуолью, цитоплазмой и клеточной оболочкой, их водные потенциалы равны. В связи с этим, как только в силу процесса транспирации возникает ненасыщенность водой клеточных стенок паренхимных клеток, она сейчас же передается внутрь клетки, водный потенциал падает, сосущая сила возрастает. Вода передвигается от клетки к клетке благодаря градиенту сосущей силы. По-видимому, передвижение воды от клетки к клетке в листовой паренхиме идет не по симпласту, а в основном по клеточным стенкам, где сопротивление значительно меньше.
Таким образом, по сосудам вода движется благодаря присасывающей силе транспирации и создающемуся в силу этого градиенту водного потенциала. Однако ни один всасывающий насос не может поднять воду на высоту больше 10 м (соответствующую 0,1 МПа нормального давления). Между тем есть деревья, у которых вода поднимается на высоту более 100 м. Объяснение этому дает теория сцепления между молекулами воды, которое в полной мере проявляется при отсутствии воздуха.
Вся вода в растении представляет единую взаимосвязанную систему. Поскольку между молекулами воды имеются силы сцепления (когезия), вода поднимается на высоту значительно большую 10 м. Сила сцепления увеличивается, так как молекулы воды обладают большим сродством друг к другу. Силы сцепления существуют и между водой и стенками сосудов. Стенки проводящих элементов ксилемы эластичны. В силу этих двух обстоятельств даже при недостатке воды связь между молекулами воды и стенками сосудов не нарушается. Это подтверждается исследованиями по изменению толщины стебля травянистых растений. Определения показали, что в полуденные часы толщина стебля травянистых растений уменьшается. Если перерезать стебель, то сосуды сразу расширяются и воздух врывается в них. Степень натяжения водных нитей в сосудах зависит от соотношения процессов поглощения и испарения воды. Все это позволяет растительному организму поддерживать единую водную систему и не обязательно восполнять каждую каплю испаряемой воды.
В том случае, если в отдельные членики сосудов попадает воздух, они, по-видимому, выключаются из общего тока проведения воды. Таков путь воды по растению и его основные движущие силы. Современные методы исследования позволяют определить скорость передвижения воды по растению. Согласно полученным данным, скорость движения воды в течение суток изменяется. В дневные часы она значительно больше. При этом разные виды растений отличаются по скорости передвижения воды. Если скорость передвижения у хвойных пород обычно не превышает 0,5-1 см/ч, то у лиственных она значительно выше. У дуба, например, скорость передвижения составляет 43,6 см/ч. Скорость передвижения воды мало зависит от напряженности обмена. Изменение температуры, введение метаболических ингибиторов не влияют на передвижение воды. Вместе с тем этот процесс, как и следовало ожидать, очень сильно зависит от скорости транспирации и от диаметра водопроводящих сосудов. В более широких сосудах вода встречает меньшее сопротивление. Однако надо учитывать, что в более широкие сосуды скорее могут попасть пузырьки воздуха или произойти какие-либо иные нарушения тока воды.
Вода поступает в растение из почвы через корневые волоски и по сосудам разносится по всей его надземной части. В вакуолях растительных клеток растворены различные вещества. Частицы этих веществ давят на протоплазму, которая хорошо пропускает воду, но препятствует прохождению через нее растворенных в воде частиц. Давление растворенных веществ на протоплазму называется осмотическим давлением. Вода, поглощенная растворенными веществами, растягивает до известного предела эластичную оболочку клетки. Как только растворенных веществ становится меньше в растворе, содержание воды уменьшается, оболочка сокращается и принимает минимальный размер. Осмотическое давление постоянно поддерживает растительную ткань в напряженном состоянии, и лишь при большой потере воды, при завядании, это напряжение - тургор - в растении прекращается.
Когда осмотическое давление уравновешено растянувшейся оболочкой, вода не может поступать в клетку. Но стоит клетке потерять часть воды, как оболочка сокращается, находящийся в клетке клеточный сок становится более концентрированным, а вода начинает поступать в клетку, пока оболочка снова не растянется и не уравновесит осмотическое давление. Чем больше воды потеряло растение, тем с большей силой вода поступает в клетки. Осмотическое давление в растительных клетках довольно велико, и его измеряют, подобно давлению в паровых котлах, атмосферами. Силу, с которой растение всасывает воду, - сосущую силу - также выражают в атмосферах. Сосущая сила у растений часто достигает 15 атмосфер и выше.
Растение непрерывно испаряет воду через находящиеся в листьях устьица. Устьица могут раскрываться и закрываться, образовывать то широкую, то узкую щель. На свету устьица раскрываются, а в темноте и при слишком большой потере воды закрываются. В зависимости от этого испарение воды идет то - интенсивно, то почти совсем прекращается.
Если срезать растение под корень, из пенька начинает сочиться сок. Это показывает, что корень и сам нагнетает воду в стебель. Следовательно, поступление воды в растение зависит не только от испарения воды через листья, но и от корневого давления. Оно перегоняет воду из живых клеток корня в полые трубки омертвевших сосудов. Так как в клетках этих сосудов нет живой протоплазмы, вода беспрепятственно движется по ним к листьям, где испаряется через устьица.
Испарение очень важно для растения. С передвигающейся водой разносятся по всему растению поглощенные корнем минеральные вещества.
Испарение снижает температуру тела растения и тем самым предохраняет его от перегрева. Растение усваивает лишь 2-3 части поглощенной им из почвы воды, остальные 997 - 998 частей испаряются в атмосферу. Чтобы образовать один грамм сухого вещества, растение в нашем климате испаряет от 300 г до килограмма воды.
Вода, поступившая в клетки корня, под влиянием разности водных потенциалов, которые возникают благодаря транспирации и корневого давления, передвигается до проводящих элементов ксилемы. Согласно современным представлениям, вода в корневой системе передвигается не только по живым клеткам. Еще в 1932г. немецкий физиолог Мюнх развил представление о существовании в корневой системе двух относительно независимых друг от друга объемов, по которым передвигается вода, - апопласта и симпласта.
Апопласт - это свободное пространство корня, в которое входят межклетные промежутки, оболочки клеток, а также сосуды ксилемы. Симпласт - это совокупность протопластов всех клеток, отграниченных полупроницаемой мембраной. Благодаря многочисленным плазмодесмам, соединяющим между собой протопласт отдельных клеток, симпласт представляет единую систему. Апопласт не непрерывен, а разделен на два объема. Первая часть апопласта расположена в коре корня до клеток эндодермы, вторая - по другую сторону клеток эндодермы и включает в себя сосуды ксилемы. Клетки эндодермы благодаря пояскам. Каспари представляют как бы барьер для передвижения воды по свободному пространству (межклетникам и клеточным оболочкам). Передвижение воды по коре корня идет главным образом по апопласту, где она встречает меньшее сопротивление, и лишь частично по симпласту.
Однако, для того, чтобы попасть в сосуды ксилемы, вода должна пройти через полупроницаемую мембрану клеток эндодермы. Таким образом, мы имеем дело как бы с осмометром, у которого полупроницаемая мембрана расположена в клетках эндодермы. Вода устремляется через эту мембрану в сторону меньшего (более отрицательного) водного потенциала. Далее вода поступает в сосуды ксилемы. Как уже упоминалось, по вопросу о причинах, вызывающих секрецию воды в сосуды ксилемы, имеются различные суждения. Согласно гипотезе Крафтса, это следствие выброса солей в сосуды ксилемы, в результате чего там создается повышенная их концентрация, и водный потенциал становится более отрицательным. Предполагается, что в результате активного (с затратой энергии) поступления соли накапливаются в клетках корня. Однако интенсивность дыхания в клетках, окружающих сосуды ксилемы (перицикла), очень низкая, и они не удерживают соли, которые благодаря этому десорбируются в сосуды. Дальнейшее передвижение воды идет по сосудистой системе корня, стебля и листа. Проводящие элементы ксилемы состоят из сосудов и трахеид.
Опыты с кольцеванием показали, что восходящий ток воды по растению движется в основном по ксилеме. В проводящих элементах ксилемы вода встречает незначительное сопротивление, что, естественно, облегчает передвижение воды на большие расстояния. Правда, некоторое количество воды передвигается и вне сосудистой системы. Однако по сравнению с ксилемой сопротивление движению воды других тканей значительно больше (не менее чем на три порядка). Это приводит к тому, что вне ксилемы движется всего от 1 до 10% общего потока воды. Из сосудов стебля вода попадает в сосуды листа. Вода движется из стебля через черешок или листовое влагалище в лист. В листовой пластинке водопроводящие сосуды расположены в жилках. Жилки, постепенно разветвляясь, становятся все более мелкими. Чем гуще сеть жилок, тем меньшее сопротивление встречает вода при передвижении к клеткам мезофилла листа. Именно поэтому густота жилкования листа считается одним из важнейших признаков ксероморфной структуры - отличительной чертой растений, устойчивых к засухе.
Иногда мелких ответвлений жилок листа так много, что они подводят воду почти к каждой клетке. Вся вода в клетке находится в равновесном состоянии. Иначе говоря, в смысле насыщенности водой имеется равновесие между вакуолью, цитоплазмой и клеточной оболочкой, их водные потенциалы равны. В связи с этим, как только в силу процесса транспирации возникает ненасыщенность водой клеточных стенок паренхимных клеток, она сейчас же передается внутрь клетки, водный потенциал которой падает. Вода передвигается от клетки к клетке благодаря градиенту водного потенциала. По-видимому, передвижение воды от клетки к клетке в листовой паренхиме идет не по симпласту, а в основном по клеточным стенкам, где сопротивление значительно меньше.
По сосудам вода движется благодаря создающемуся в силу транспирации градиенту водного потенциала, градиенту свободной энергии (от системы с большей свободой энергии к системе с меньшей). Можно привести примерное распределение водных потенциалов, которое и вызывает передвижение воды: водный потенциал почвы (0,5 бара), корня (2 бара), стебля (5 бар), листьев (15 бар), воздуха при относительной влажности 50% (1000 бар).
Однако ни один всасывающий насос не может поднять воду на высоту больше 10м. Между тем есть деревья, у которых вода поднимается на высоту более 100м. Объяснение этому дает теория сцепления, выдвинутая русским ученым Е. Ф. Вотчалом и английским физиологом Е. Диксоном. Для лучшего понимания рассмотрим следующий опыт. В чашку с ртутью помещают заполненную водой трубку, которая заканчивается воронкой из пористого фарфора. Вся система лишена пузырьков воздуха. По мере испарения воды ртуть поднимается по трубке. При этом высота подъема ртути превышает 760мм. Это объясняется наличием сил сцепления между молекулами воды и ртути, которые в полной мере проявляются при отсутствии воздуха. Сходное положение, только еще более ярко выраженное, имеется в сосудах у растений.
Вся вода в растении представляет единую взаимосвязанную систему. Поскольку между молекулами воды имеются силы сцепления (когезия), вода поднимается на высоту, значительно большую 10м. Расчеты показали, что благодаря наличию сродства между молекулами воды силы сцепления достигают величины - 30 бар. Это такая сила, которая позволяет поднять воду на высоту, равную 120м, без разрыва водных нитей, что примерно и составляет максимальную высоту деревьев. 120м, без разрыва водных нитей, что примерно и составляет максимальную высоту деревьев. Силы сцепления существуют и между водой и стенками сосудов (адгезия). Стенки проводящих элементов ксилемы эластичны. В силу этих двух обстоятельств даже при недостатке воды связь между молекулами воды и стенками сосудов не нарушается.